与传统抗体相比,重组抗体具有几个关键优势。这些包括良好的批次间一致性,持续供应以及对抗体工程的适应性。因此,重组抗体在科学研究中的应用日益广泛,特别是作为解决持续存在的可重复性难题的一种手段。

传统的多克隆和单克隆抗体是正常 B 细胞发育和基因重组的产物。它们是通过用抗原免疫动物以引发免疫应答而产生的。多克隆抗体由许多不同的 B 细胞克隆分泌并识别多个抗原表位,而单克隆抗体则来自单个 B 细胞克隆,并且仅对单个表位具有特异性。
重组抗体是单克隆抗体,但是其生产涉及体外基因操作。将抗体基因克隆到表达载体中后,将其转染到合适的宿主细胞系中进行抗体表达。哺乳动物细胞系最常用于重组抗体的生产,然而细菌、酵母或昆虫来源的细胞系也适用。
由于重组抗体的生产涉及对抗体轻链和重链进行测序,因此这是一个高度可控且可靠的过程。相反,用于生产单克隆抗体的基于杂交瘤的系统容易发生遗传漂移和不稳定,从而增加了批次间变异或抗体表达缺失的可能性。重组抗体在批次之间高度一致,从而确保了可重复的实验结果。
体外生产抗体的方法适合大规模生产,这意味着抗体的可获得性不太可能成为限制因素。此外,由于重组抗体序列是已知的,因此确保了供应的连续性;如需将抗体用于大规模长期研究,这可能就是一个至关重要的因素。
与传统的抗体生产方法不同,重组方法避免了使用动物的需要。多克隆抗体直接从免疫宿主血清中纯化,单克隆抗体从杂交瘤来源的组织培养上清液 (TCS) 或腹水中纯化,而重组抗体是从转染宿主细胞系的 TCS 中纯化。无论抗体是多克隆抗体、单克隆抗体还是重组抗体,在实验使用前都必须在预期应用中对其进行适当验证。在CST,我们遵守抗体验证标志,即在任何特定实验方法中的确定抗体特异性、敏感性和功能性的六个互补策略。通过针对每种抗体产品精心定制这些策略,我们保证 CST 抗体的适用性,以帮助您获得可信赖的结果。

| 反应性 | H M R |
| 灵敏度 | 内源性 |
| MW (kDa) | 62 |
| 来源/同种型 | 兔 IgG |
产品信息
人, 小鼠, 大鼠
仓鼠 , 猴, 牛 , 猪 , 马
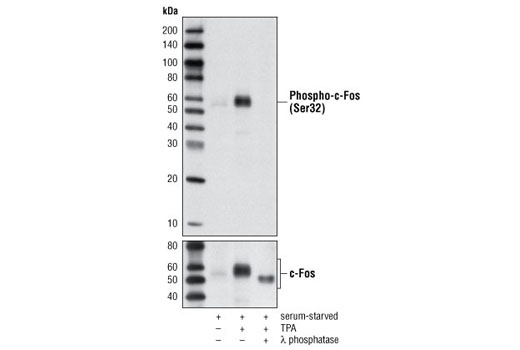
使用与人 c-Fos 蛋白的 Ser32 周围残基相对应的合成磷酸肽,对动物进行免疫接种来产生单克隆抗体。
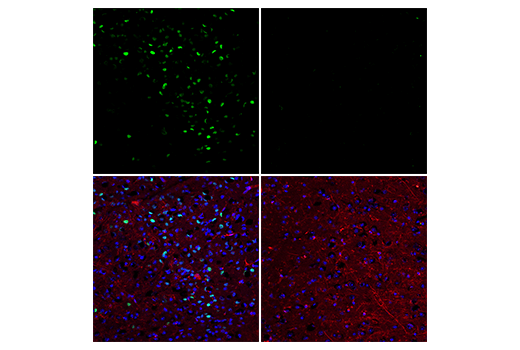
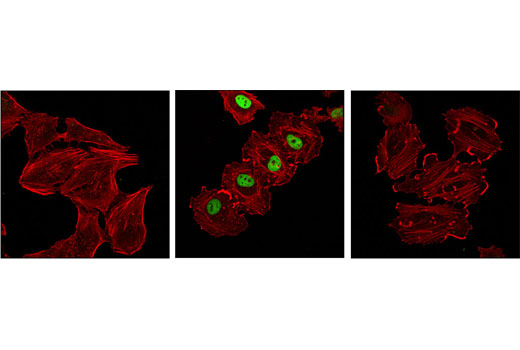
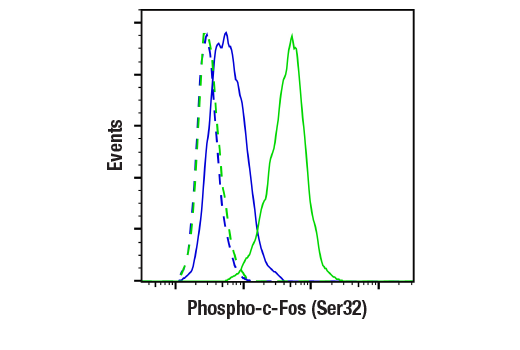
胞核癌基因 Fos 家族包括 c-Fos、FosB、Fos-相关抗原 1 (FRA1) 和 Fos-相关抗原 2 (FRA2) (1)。虽然大部分 Fos 蛋白作为单一亚型存在,但是 FosB 蛋白作为两种亚型存在:全长 FosB 和缩短形式 FosB2 (δ FosB),后者缺乏羧基端的 101 个氨基酸 (1-3)。Fos 蛋白的表达可由各种胞外刺激快速且瞬间诱导,这些刺激包括生长因子、细胞因子、神经递质、多肽激素和应激反应。Fos 蛋白与 Jun 蛋白(c-Jun、JunB 和 JunD)二聚化以形成激活蛋白-1 (AP-1),一种与 TRE/AP-1 元件结合并激活转录的转录因子。Fos 蛋白和 Jun 蛋白包含可介导二聚化的亮氨酸拉链基序和一个与 DNA 结合的毗邻碱性结构域。各种 Fos/Jun 异二聚体在反式激活 AP-1 依赖性基因方面的能力不同。Erk 激酶引起的 Fos 蛋白磷酸化对胞外刺激应答后,除了表达增加外,还可能会进一步增强转录活性 (4-6)。Erk5 引起的 c-Fos 在 Ser32 和 Thr232 的磷酸化增强了蛋白质稳定性及核定位 (5)。Erk1/2 引起的 FRA1 在 Ser252 和 Ser265 的磷酸化增强了蛋白质稳定性并导致 FRA1 在癌细胞中过量表达 (6)。生长因子刺激后,FosB 和 c-Fos 的表达在静息成纤维细胞中立即开始,但是持续时间很短,蛋白水平在几小时后便消失了 (7)。FRA1 和 FRA2 表达持续的时间更长,并且可以在非同步生长的细胞中检测到它的相对水平 (8)。去调控的 c-Fos、FosB 和 FRA2 表达可能导致肿瘤性细胞转化;但 δ FosB 不能转化细胞 (2,3)。
探索与本品相关的通路。
STRING - 已知和预测的蛋白质间相互作用。
除非如以 CST 合法授权代表签署的书面形式另行明确同意,否则以下条款适用于 CST、其附属公司或其分销商提供的产品。除非 CST 合法授权代表以书面形式单独接受,否则任何附加于或异于此处所载条款和条件的客户条款和条件均被拒绝且无效。
产品用“仅供研究使用”或类似标示声明标示,并且尚未经 FDA 或其他国外或国内监管实体出于任何目的批准、准许或许可。客户不得出于任何诊断或治疗目的或以任何与产品标示声明相冲突的方式使用任何产品。CST 销售或许可的产品提供给作为最终用户的客户,且仅用于研究和开发用途。出于诊断、预防或治疗目的任何产品使用或出于转售(单独或作为成分)或其他商业目的的任何产品购买都要求来自 CST 的单独许可。客户 (a) 不得向任何第三方出售、许可、出借、捐赠或另行转让或提供任何本公司产品,无论单独或联合其他材料方式,或使用本公司产品制造任何商业产品,(b) 不得复制、修改、逆向工程、反编译、反汇编或另行尝试发现本公司产品的底层结构或技术,或出于开发与 CST 产品或服务竞争的任何产品或服务的目的使用本公司产品,(c) 不得从本公司产品改变或移除任何商标、商品名称、徽标、专利或版权声明或标记,(d) 仅应根据 CST 产品销售条款和任何适用文档使用本公司产品,以及 (e) 应就客户联系本公司产品所用的任何第三方产品或服务而言遵守任何许可、服务条款或类似协议。